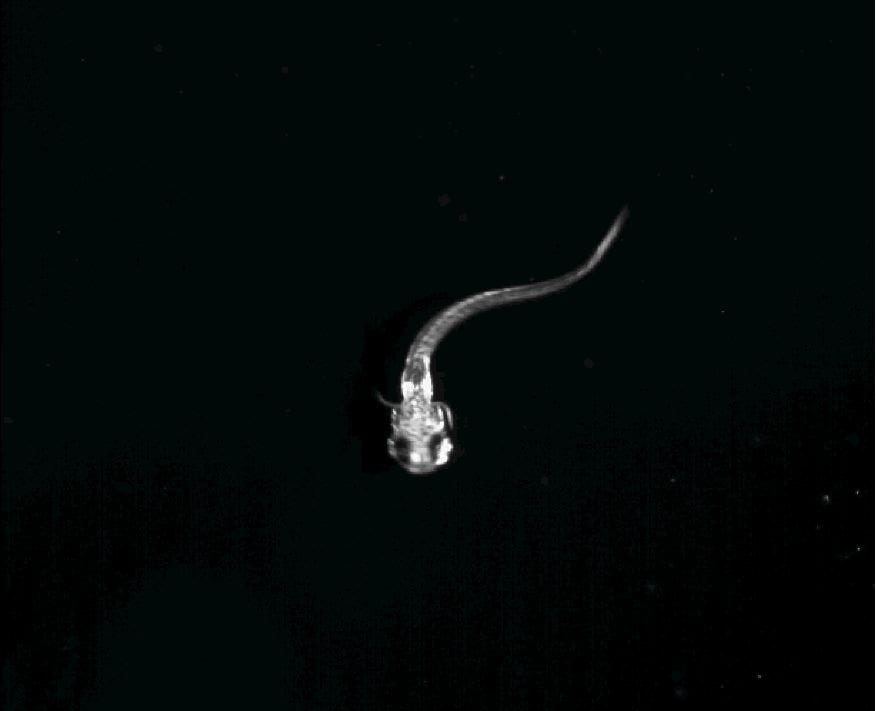
LAUDER LABORATORY George V. Lauder Research Areas Featured Video Robotic Maneuver Photron Pectoral Fin Ventral Learn About the Lab People Research